Which are less readily detected by conventional mass spectrometry approaches, and to keep the database to a reasonable size. Finally, we used the NCBI reference sequence database as a source for RNA transcripts. This non-redundant and well-annotated database is fairly conservative, and thus is a quality source for identifying candidate AltORFs, but it would be interesting to compare with other databases to verify if different mRNA isoforms could also serve as template for the expression of corresponding alternative proteins. Nevertheless, the location of AltORfs is comparable with the distribution obtained in a peptidomic study of small ORFs encoded polypeptides, with the exception of AltORFs located in 59UTRs. For these AltORFs, the apparent discrepancy probably results from our prediction of AltORFs initiating at AUG sites only. In conclusion, we have provided compelling evidence that alternative proteins significantly contribute to the human proteome by identifying 1259 new proteins and many more will likely be detected in further MS experiments. A comprehensive knowledge of the proteome is of crucial interest to unravel the cellular mechanisms underlying KRX-0401 supply health and disease. We believe that proteomics approaches supported by ribosome profiling will further benefit the establishment of an exhaustive catalog of proteins to fulfill this goal in the future. Higher organisms are continuously exposed to a great variety of pathogens such as viruses, mycoplasma, bacteria, and fungi. To fight these microbes they have developed several defense strategies, including the production of antimicrobial peptides. AMPs are effective weapons against a wide range of pathogens and are distributed throughout the animal and plant kingdom, suggesting that they are critical for the successful evolution of complex multicellular organisms. Despite their high sequence diversity, AMPs share fundamental structural properties such as short size, positive net charge, hydrophobic nature and clustering of cationic and hydrophobic amino acids within distinct domains of the molecule. Upon contact with pathogen membranes AMPs tend to adopt amphiphilic structures. Because of their cationic and hydrophobic features, antimicrobial peptides interact primarily with negatively charged biomembranes. Many bacterial membranes contain negatively charged components like hydroxylated phospholipids, lipopolysaccharides and teichonic acids and are therefore major targets for AMPs. The hydrophobic regions of the AMPs support incorporation of the peptides into the membranes, leading to pore formation and permeabilization. Several different models have been proposed for peptide insertion, of which the barrel-stave model, the carpet model, and the toroidal-pore model are the most popular ones. In plant protection, bacterial infections are hard to overcome, considering that plant disease control is mainly based on the application of chemical pesticides, which are under strong restrictions and regulatory requirements. However, about 14% of the total loss of all crops produced worldwide are caused by infectious diseases, resulting in a total annual loss of about 220 $ billion per year �C not including the 6�C12% losses of crop after harvest. Furthermore, microbial organisms often produce toxic compounds, which make food products uneatable or even dangerous for humans and animals. Therefore there is an urgent need for new antimicrobial agents. AMPs have attracted the interest of researchers for many years. Especially their mode of action, namely targeting  fundamental features of microbial cell membranes, is thought to reduce the risk of resistance development in microbial populations �C as it happened in the past to every new antibiotic within a few years of its utilization since this would Nutlin-3 require a reorganization of the bacterial membranes.
fundamental features of microbial cell membranes, is thought to reduce the risk of resistance development in microbial populations �C as it happened in the past to every new antibiotic within a few years of its utilization since this would Nutlin-3 require a reorganization of the bacterial membranes.
We introduced a cut-off of amino acids to discard from our databases polypeptides shorter than residues
Leave a reply
 HspB1 amino terminus, which is involved in phosphorylation sensitive interactions, is crucial for oligomerization. Consequently, HspB1 and HspB5 holdase chaperone activity are regulated by the complex relationship that exists between their phosphorylation and oligomerization status. For example, it is particularly intriguing to note that in cells exposed to different environmental conditions or insults, HspB1 displays stress-specific changes in its oligomerization/phosphorylation status. Consequently, HspB1 probably acts as a protein sensor, which through structural changes, can interact with the most appropriate client protein targets. These phenomena subsequently allow cells to adapt to changes in their environment and/or mount a protective anti-stress response. In tissues that express several sHsps, such as in lens and
HspB1 amino terminus, which is involved in phosphorylation sensitive interactions, is crucial for oligomerization. Consequently, HspB1 and HspB5 holdase chaperone activity are regulated by the complex relationship that exists between their phosphorylation and oligomerization status. For example, it is particularly intriguing to note that in cells exposed to different environmental conditions or insults, HspB1 displays stress-specific changes in its oligomerization/phosphorylation status. Consequently, HspB1 probably acts as a protein sensor, which through structural changes, can interact with the most appropriate client protein targets. These phenomena subsequently allow cells to adapt to changes in their environment and/or mount a protective anti-stress response. In tissues that express several sHsps, such as in lens and  It was proposed that Tic22 is processed in the intermembrane space by an unknown peptidase, and that its targeting pathway is different from other known routes. These observations may account for our failure to observe efficient chloroplast targeting of a range of different full-length and truncated Tic22 fusions to YFP in transfected protoplasts. It is conceivable that this unusual import pathway is unable to accommodate such heterologous passenger proteins, or that the C-terminal addition of a large tag like YFP somehow disrupts the targeting signals of the Tic22 protein. Regardless, there is little doubt that the Arabidopsis Tic22 proteins are chloroplast localized, as this has been shown by in vitro import, protease treatment of isolated chloroplasts, and by the proteomic analysis of purified chloroplast fractions. The data presented in this report complement those of Rudolf et al. in the following ways: we conducted a comprehensive analysis of the evolution of the Tic22 gene family; we identified and characterized a tic22-III single mutant phenotype; we identified and characterized more than one tic22 double-mutant genotype; we analysed the accumulation of a range of different chloroplast proteins in the tic22 mutants by immunoblotting; we conducted a quantitative analysis of chloroplasts using electron microscopy in single as well as double-mutant plants.
It was proposed that Tic22 is processed in the intermembrane space by an unknown peptidase, and that its targeting pathway is different from other known routes. These observations may account for our failure to observe efficient chloroplast targeting of a range of different full-length and truncated Tic22 fusions to YFP in transfected protoplasts. It is conceivable that this unusual import pathway is unable to accommodate such heterologous passenger proteins, or that the C-terminal addition of a large tag like YFP somehow disrupts the targeting signals of the Tic22 protein. Regardless, there is little doubt that the Arabidopsis Tic22 proteins are chloroplast localized, as this has been shown by in vitro import, protease treatment of isolated chloroplasts, and by the proteomic analysis of purified chloroplast fractions. The data presented in this report complement those of Rudolf et al. in the following ways: we conducted a comprehensive analysis of the evolution of the Tic22 gene family; we identified and characterized a tic22-III single mutant phenotype; we identified and characterized more than one tic22 double-mutant genotype; we analysed the accumulation of a range of different chloroplast proteins in the tic22 mutants by immunoblotting; we conducted a quantitative analysis of chloroplasts using electron microscopy in single as well as double-mutant plants. LEPR, many other genes including CytokineR, JAK, STAT, downstream genes and regulatory molecules are first fully and systemically identified in crab. Considering different aspects of cell development and host response activated by JAK-STAT pathway, there is no surprise that lots of regulators are found to control this pathway. Expression and regulation of components in JAK-STAT pathway are also reported in transcriptome analyses of microbial infected Pseudosciaena crocea and Laodelphax striatellus.
LEPR, many other genes including CytokineR, JAK, STAT, downstream genes and regulatory molecules are first fully and systemically identified in crab. Considering different aspects of cell development and host response activated by JAK-STAT pathway, there is no surprise that lots of regulators are found to control this pathway. Expression and regulation of components in JAK-STAT pathway are also reported in transcriptome analyses of microbial infected Pseudosciaena crocea and Laodelphax striatellus.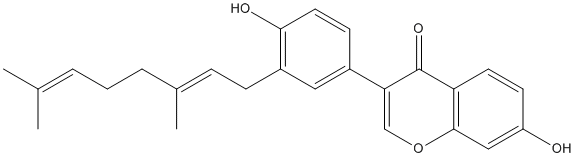 KO, as contrasted with antidepressant-like phenotype of the NET and DAT KO mice. Thus, the mainly frontal MEMRI tracing in DAT and NET KO mice is likely to be indicative of more robust activity and/or connectivity in the frontal portion of the reward circuit of the DAT and NET KO mice compared to wildtype or the SERT KO mice. In the context of understanding the genetic basis of mental illness, Robbins and Arnsten review monoaminergic influences on executive functions of the PFC in rodents, nonhuman primates and humans. They note that much subtler genetic alterations in norepinephrine and dopamine signaling may have significant influences on susceptibility for major mental illnesses. Therefore findings about these mixtures are most often attributed to CUR. CUR belongs to the pharmacopoeia of Asian traditional medicine or alternative medicine to treat inflammatory diseases and a wide range of disorders. Given the role of inflammation in the promotion of chronic human diseases including Alzheimer’s disease, chronic obstructive pulmonary disease, cataract, diabetes, and cancer, CUR has deserved extensive research.
KO, as contrasted with antidepressant-like phenotype of the NET and DAT KO mice. Thus, the mainly frontal MEMRI tracing in DAT and NET KO mice is likely to be indicative of more robust activity and/or connectivity in the frontal portion of the reward circuit of the DAT and NET KO mice compared to wildtype or the SERT KO mice. In the context of understanding the genetic basis of mental illness, Robbins and Arnsten review monoaminergic influences on executive functions of the PFC in rodents, nonhuman primates and humans. They note that much subtler genetic alterations in norepinephrine and dopamine signaling may have significant influences on susceptibility for major mental illnesses. Therefore findings about these mixtures are most often attributed to CUR. CUR belongs to the pharmacopoeia of Asian traditional medicine or alternative medicine to treat inflammatory diseases and a wide range of disorders. Given the role of inflammation in the promotion of chronic human diseases including Alzheimer’s disease, chronic obstructive pulmonary disease, cataract, diabetes, and cancer, CUR has deserved extensive research.