After each heating cycle, the activity of individual enzyme molecules changed randomly from its prior activity, with approximately 50% of the population gaining activity and 50% losing activity. Considering the error associated with linear fitting, a maximum of 12% of the enzyme population may have Tetrahydroberberine overlapping activity values after each pulse. There are several possible results that may be expected to occur upon heating an enzyme. First, if the activation barrier is too high for the molecule to convert to a different local minimum, the enzyme will fall back into the same local minimum, maintain its original conformation, and there will be no change in the reaction rate after heating. Second, if the enzyme overcomes the energy barrier, it may convert to a new stable conformation. The new conformation may have either a faster or slower reaction rate. The changes in activity could also be a result of the tetramer-dimermonomer equilibrium. At the single molecule level, however, the tetramer-dimer-monomer equilibrium is shifted to the monomer form due to the low concentration of monomer in the microwells such that if the tetramer dissociates, we would expect an irreversible loss in enzyme activity��a phenomenon we observe for a small number of enzyme molecules in these experiments. Narrowing of the distribution after the first heating pulse may be caused by kinetic traps that have slightly higher energy barriers and are only accessible when the nascent protein folds; thus some enzymes cannot return to their previous states upon re-heating at 47uC. No correlation between the activities of enzymes before and after heating was observed, suggesting that enzymes have no ‘memory’ of any previous 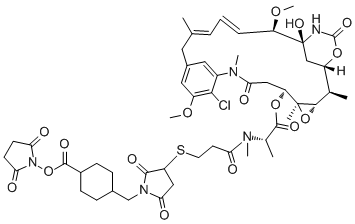 conformations before the heating pulse was introduced. The change in activity of each enzyme was random as well, i.e. it did not correlate with its initial activity. Each enzyme exhibits an equal probability of either gaining or losing activity through all heating periods. As temperature increases, a protein molecule is expected to gain enough energy to overcome an energy barrier and convert to a new conformation that has a different activity. If conformational differences between different enzyme molecules are the cause of the broad activity distribution, then the activities of the individual molecules should redistribute after each heating pulse, which is what we observe. On the other hand, if sequence differences are the sole basis for the activity distribution, then upon heating, each enzyme molecule should revert to its original activity. Our results clearly show that virtually all the enzyme molecules redistribute their activity when heated, demonstrating the importance of conformations in static heterogeneity. It is important to note that both conformation and sequence may contribute to the activity distribution because enzyme molecules with different sequences will also interconvert between conformations upon heating. Loganin Moreover, enzymes with different primary sequences may display varying kinetic responses upon heating. In addition to the observed change in activity due to conformational changes, denaturation of some enzyme molecules occurred, which is likely the reason for some loss in activity when activity is measured in bulk solution. These enzymes are irreversibly denatured and contribute to the decrease in average activity over time. The denaturation temperature of b-galactosidase was calculated to be 55uC by CD experiments and confirmed by kinetic measurements, and agrees well with denaturation temperatures reported in the literature. In the present experiments, denaturation of the enzymes was minimized by heating only to 47uC, which is below this denaturation temperature.
conformations before the heating pulse was introduced. The change in activity of each enzyme was random as well, i.e. it did not correlate with its initial activity. Each enzyme exhibits an equal probability of either gaining or losing activity through all heating periods. As temperature increases, a protein molecule is expected to gain enough energy to overcome an energy barrier and convert to a new conformation that has a different activity. If conformational differences between different enzyme molecules are the cause of the broad activity distribution, then the activities of the individual molecules should redistribute after each heating pulse, which is what we observe. On the other hand, if sequence differences are the sole basis for the activity distribution, then upon heating, each enzyme molecule should revert to its original activity. Our results clearly show that virtually all the enzyme molecules redistribute their activity when heated, demonstrating the importance of conformations in static heterogeneity. It is important to note that both conformation and sequence may contribute to the activity distribution because enzyme molecules with different sequences will also interconvert between conformations upon heating. Loganin Moreover, enzymes with different primary sequences may display varying kinetic responses upon heating. In addition to the observed change in activity due to conformational changes, denaturation of some enzyme molecules occurred, which is likely the reason for some loss in activity when activity is measured in bulk solution. These enzymes are irreversibly denatured and contribute to the decrease in average activity over time. The denaturation temperature of b-galactosidase was calculated to be 55uC by CD experiments and confirmed by kinetic measurements, and agrees well with denaturation temperatures reported in the literature. In the present experiments, denaturation of the enzymes was minimized by heating only to 47uC, which is below this denaturation temperature.
Enzymes that did not show any activity after three consecutive heating pulses were considered denatured
Leave a reply
 and short-looped nephrons, which are classified according to their long and short-looped MALs. The functional differences between lMALs and sMALs are not well known. The proportion of lMALs and sMALs differs among animals. Humans have a larger number of sMALs than lMALs. In contrast, rats and mice have a larger number of lMALs than sMALs. The pocket mouse has a 10-fold higher single-nephron glomerular filtration rate via long-looped nephrons compared with short-looped nephrons. We have previously shown that AVP-stimulated NaCl
and short-looped nephrons, which are classified according to their long and short-looped MALs. The functional differences between lMALs and sMALs are not well known. The proportion of lMALs and sMALs differs among animals. Humans have a larger number of sMALs than lMALs. In contrast, rats and mice have a larger number of lMALs than sMALs. The pocket mouse has a 10-fold higher single-nephron glomerular filtration rate via long-looped nephrons compared with short-looped nephrons. We have previously shown that AVP-stimulated NaCl  of cell cycle. Several studies have shown that BTG3 was associated with tumorigenesis. It has been reported as a candidate tumor suppressor gene in several cancers. However, the expression pattern and functions of BTG3 in the progression of HCC remain elusive. In this study, we provide evidence that BTG3 plays an important suppressive role in HCC progression especially tumor cell proliferation, invasion and cell cycle progression. First, we detected the expression of BTG3 in a large cohort of clinical paraffin-embedded HCC tissues with long-term followup, 20 paired fresh HCC tissues and 5 HCC cell lines. The results showed that BTG3 had a higher probability of being down-regulated in HCC tissues. BTG3 expression was an independent prognostic marker for survival of HCC patients. Moreover, BTG3 expression, portal vein thrombosis, differentiation, distant metastasis, dissemination and relapse were singled out as six marked and independent prognostic factors relative to overall survival. Besides BTG3 expression, the five other factors are
of cell cycle. Several studies have shown that BTG3 was associated with tumorigenesis. It has been reported as a candidate tumor suppressor gene in several cancers. However, the expression pattern and functions of BTG3 in the progression of HCC remain elusive. In this study, we provide evidence that BTG3 plays an important suppressive role in HCC progression especially tumor cell proliferation, invasion and cell cycle progression. First, we detected the expression of BTG3 in a large cohort of clinical paraffin-embedded HCC tissues with long-term followup, 20 paired fresh HCC tissues and 5 HCC cell lines. The results showed that BTG3 had a higher probability of being down-regulated in HCC tissues. BTG3 expression was an independent prognostic marker for survival of HCC patients. Moreover, BTG3 expression, portal vein thrombosis, differentiation, distant metastasis, dissemination and relapse were singled out as six marked and independent prognostic factors relative to overall survival. Besides BTG3 expression, the five other factors are  aberrant roles of BTG3 in HCC progression. Second, a series of relevant functional experiments in vitro, from positive to negative, were performed in our study. Here, we observed that BTG3 could strongly inhibit the proliferative abilities of HCC cell lines. As a member of the anti-proliferative gene family, over-expression of BTG3 also inhibits cell proliferation in breast cancer. The inhibition of cell proliferation by BTG3 is thought to result from its negative regulation of cell cycle. Our flow cytometric analysis showed that BTG3 was expressed highly in late G1 phase before the entry of the cells in S phase, while down-regulation of BTG3 promoted G1/S cycle transition of HCC cells. Several reports demonstrate that BTG3 constitutes important negative regulatory mechanism for Src-mediated signaling and it inhibits transcription factor E2F1, suggesting it has a negative regulatory influence consistent with its role to inhibit progression into S-phase, meanwhile, BTG3 was identified as a transcriptional target of p53. BTG3 interacts with CHK1, a key effector kinase in the cell cycle checkpoint response, and regulates its phosphorylation and activation. Moreover, MiR-378
aberrant roles of BTG3 in HCC progression. Second, a series of relevant functional experiments in vitro, from positive to negative, were performed in our study. Here, we observed that BTG3 could strongly inhibit the proliferative abilities of HCC cell lines. As a member of the anti-proliferative gene family, over-expression of BTG3 also inhibits cell proliferation in breast cancer. The inhibition of cell proliferation by BTG3 is thought to result from its negative regulation of cell cycle. Our flow cytometric analysis showed that BTG3 was expressed highly in late G1 phase before the entry of the cells in S phase, while down-regulation of BTG3 promoted G1/S cycle transition of HCC cells. Several reports demonstrate that BTG3 constitutes important negative regulatory mechanism for Src-mediated signaling and it inhibits transcription factor E2F1, suggesting it has a negative regulatory influence consistent with its role to inhibit progression into S-phase, meanwhile, BTG3 was identified as a transcriptional target of p53. BTG3 interacts with CHK1, a key effector kinase in the cell cycle checkpoint response, and regulates its phosphorylation and activation. Moreover, MiR-378