In this respect, it is interesting to note that the heating rate explained more than 78% of the variation in Bcl2:Bax transcripts in A. millepora during our thermal stress experiments. This result suggests that the heating rate could be an important environmental driver of this molecular response in the host rather than duration and intensity of thermal anomalies. However, because the thermal treatments used in this study varied in both the average temperature and the heating rate, the effects observed on the level of expression of Bcl-2 family members are attributable to some combination of these 2 factors. Future studies should therefore include thermal treatments with tight control of temperature in order to precise the relative contributions of each factor on the molecular regulation of apoptosis in corals. This study provides further evidence that Tulathromycin B corals can regulate apoptosis in response to thermal stress. Bleaching is nevertheless an extremely dynamic event with many different triggers, responses and consequences to coral-dinoflagellate symbiosis. Understanding the cellular dynamics and the mechanisms driving the breakdown of this symbiosis will require many further studies but this is a critical step to predict future bleaching events and coral mortality under a changing climate. The formation of embryonic tissues is a key feature in generating diversity in animal development. After cell fate is established, cell-cell signaling and intracellular signal transduction pathways instruct cells to undergo cell shape changes. These cell shape changes are necessary for cell movement, a basic process that underlies embryonic development and is largely accomplished by regulation of the actin cytoskeleton. Actin dynamics is required for the migration of individual groups of cells, as in border cell migration in the Drosophila ovary, or large groups of cells, such as those involved in gastrulation in the developing fly embryo. One common feature of cell rearrangements via the actin cytoskeleton is the involvement of the Rho family of GTPases. Widely conserved across species and involved in seemingly diverse developmental processes including cell migration, phagocytosis, and myoblast fusion, the Rho GTPases are key signaling molecules that impinge upon actin cytoskeletal reorganization. Several classes of GTPase regulatory proteins have been identified, including the GTPase-activating proteins, guanine nucleotide exchange factors, and guanine nucleotide dissociation inhibitors. In particular, the GEFs regulate GTPase activity by exchanging the inactive, GDP-bound Rac to the active, GTP-bound state. It is thought that GEFs are a crucial intermediate that signal from upstream cell surface receptors to mediate GTPase activation. Some GEFs directly associate with membrane receptors, while others are associated via an intermediate complex. In flies, two neuronally expressed Rac GEFs have been identified that exemplify this in development of the central nervous system. Trio physically interacts with the Netrin receptor Frazzled to regulate chemoattraction, while Son of sevenless associates with the Roundabout receptor through the SH2-SH3 adaptor protein Dreadlocks to control axon repulsion. Vertebrate cell culture studies show this region is required for membrane localization. In flies, the DHR1 domain ![]() is not essential for recruitment to the membrane, but is essential for myoblast fusion as deletion of the DHR1 domain fails to rescue mbc mutant embryos in functional rescue assays. Although the SH3-domain containing protein Crk is capable of binding the C-terminal proline-rich region of both DOCK180 and Mbc, it is not 3,4,5-Trimethoxyphenylacetic acid always essential in vivo. A direct interaction between vertebrate DOCK180 and CrkII is not required for apoptotic cell removal. Conventional GEFs bind nucleotide-free Rac via their DH domain, while the CDM proteins use the DHR2 region. Deletion or mutation of this domain results in a loss of Rac binding and activation. A DOCK-Rac protein complex is sufficient for Rac activation, but may be enhanced by DOCK180 bound to ELMO.
is not essential for recruitment to the membrane, but is essential for myoblast fusion as deletion of the DHR1 domain fails to rescue mbc mutant embryos in functional rescue assays. Although the SH3-domain containing protein Crk is capable of binding the C-terminal proline-rich region of both DOCK180 and Mbc, it is not 3,4,5-Trimethoxyphenylacetic acid always essential in vivo. A direct interaction between vertebrate DOCK180 and CrkII is not required for apoptotic cell removal. Conventional GEFs bind nucleotide-free Rac via their DH domain, while the CDM proteins use the DHR2 region. Deletion or mutation of this domain results in a loss of Rac binding and activation. A DOCK-Rac protein complex is sufficient for Rac activation, but may be enhanced by DOCK180 bound to ELMO.
Importantly studies in Drosophila have provided additional insight into mechanisms in corals exposed to sudden environmental changes
Leave a reply
 have been proposed to explain DHA’s anti-apoptotic and anti-inflammatory effects, including maintenance of plasma membrane integrity, activation of Akt signaling, and conversion into other derivatives. These findings also provide clues for NPD1’s potential targets. NPD1 inhibits NFkB activation and COX-2 expression in brain ischemia-reperfusion, while Ab peptide-induced apoptosis is associated with ERK and p38 MAPK-NF-kB mediated COX-2 up-regulation. Neuroprotection mediated by NPD1 may further involve components of signaling pathways upstream of NF-kB activation and DNA-binding. Our results provide compelling evidence that NPD1 is endowed with strong anti-inflammatory, anti-amyloidogenic, and antiapoptotic bioactivities in HNG cells upon exposure to Ab42 oligomers, or in HNG cells over-expressing bAPPsw. These results suggest that NPD1’s anti-amyloidogenic effects are mediated in part through activation of the PPARc receptor, while NPD1’s stimulation of non-amyloidogenic pathways is PPARc-independent. Suggested sites of NPD1 actions are schematically presented in Figure 11. NPD1 stimulation of ADAM10 coupled to suppression of BACE1-mediated Ab42 secretion clearly warrants further study, as these dual secretase-mediated pathways may provide effective combinatorial or multi-target approaches in the clinical management of the AD process. Tau is an axonally located, microtubule-associated protein that is encoded by a single gene and predominantly expressed in neurons. Tau mRNA transcripts can be spliced alternatively, and the expression of tau-isoforms is developmentally regulated and varies between species.
have been proposed to explain DHA’s anti-apoptotic and anti-inflammatory effects, including maintenance of plasma membrane integrity, activation of Akt signaling, and conversion into other derivatives. These findings also provide clues for NPD1’s potential targets. NPD1 inhibits NFkB activation and COX-2 expression in brain ischemia-reperfusion, while Ab peptide-induced apoptosis is associated with ERK and p38 MAPK-NF-kB mediated COX-2 up-regulation. Neuroprotection mediated by NPD1 may further involve components of signaling pathways upstream of NF-kB activation and DNA-binding. Our results provide compelling evidence that NPD1 is endowed with strong anti-inflammatory, anti-amyloidogenic, and antiapoptotic bioactivities in HNG cells upon exposure to Ab42 oligomers, or in HNG cells over-expressing bAPPsw. These results suggest that NPD1’s anti-amyloidogenic effects are mediated in part through activation of the PPARc receptor, while NPD1’s stimulation of non-amyloidogenic pathways is PPARc-independent. Suggested sites of NPD1 actions are schematically presented in Figure 11. NPD1 stimulation of ADAM10 coupled to suppression of BACE1-mediated Ab42 secretion clearly warrants further study, as these dual secretase-mediated pathways may provide effective combinatorial or multi-target approaches in the clinical management of the AD process. Tau is an axonally located, microtubule-associated protein that is encoded by a single gene and predominantly expressed in neurons. Tau mRNA transcripts can be spliced alternatively, and the expression of tau-isoforms is developmentally regulated and varies between species.-Pugnac-chemical-structure.gif) in differential modes of receptor regulation. At low concentrations of xanomeline, down-regulation or internalization may be the predominant mechanism occurring to explain the appearance of a high-potency phase of
in differential modes of receptor regulation. At low concentrations of xanomeline, down-regulation or internalization may be the predominant mechanism occurring to explain the appearance of a high-potency phase of 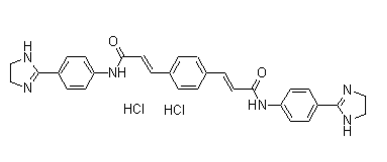 both the cellular and embryonic level. Consistent with a role in such a pathway, phenotypes in Hipk1 morphants closely resembled those observed in embryos over-expressing Hipk1, including comparisons between dorsal versus ventral injections. When either Hipk1MO was injected into the DMZ, phenotypes included shortened embryos with defects in blastopore closure and in neural tube closure. Dsh is a critical signaling molecule involved in many Wntrelated activities during gastrulation including cell fate determination, cell shape, and cell movement. Although Dsh is expressed fairly ubiquitously, it exhibits variable intracellular localization, signaling activities, and protein interactions over the course of early X. laevis development. Consequently, it is important to identify binding partners of Dsh that mediate alternate developmental functions. In this study we have identified one such protein as the nuclear kinase Hipk1, and have shown that it also can interact with the Wnt/b-catenin transcriptional corepressor Tcf3. Proper germ layer specification reflected by
both the cellular and embryonic level. Consistent with a role in such a pathway, phenotypes in Hipk1 morphants closely resembled those observed in embryos over-expressing Hipk1, including comparisons between dorsal versus ventral injections. When either Hipk1MO was injected into the DMZ, phenotypes included shortened embryos with defects in blastopore closure and in neural tube closure. Dsh is a critical signaling molecule involved in many Wntrelated activities during gastrulation including cell fate determination, cell shape, and cell movement. Although Dsh is expressed fairly ubiquitously, it exhibits variable intracellular localization, signaling activities, and protein interactions over the course of early X. laevis development. Consequently, it is important to identify binding partners of Dsh that mediate alternate developmental functions. In this study we have identified one such protein as the nuclear kinase Hipk1, and have shown that it also can interact with the Wnt/b-catenin transcriptional corepressor Tcf3. Proper germ layer specification reflected by