Finding clear evidence for disrupted coordination of mitochondrial gene expression led us to carryout a global analysis of skeletal Diperodon muscle phenotype using microarray technology to determine the extent of altered muscle phenotype. Informatic analysis yielded profound evidence for degeneration and loss of muscle specific genes. We found that regulation of apoptosis, proteosome function, ion homeostasis and kinase signalling were modulated and these appeared to us as rather predictable findings. Gene ontology analysis of the down regulated list indicated that there was a dramatic loss of unique muscle related gene expression demonstrating that the patient muscle tissue was undergoing a dedifferentiation process. Down regulated genes also indicated a loss of expression of numerous extracellular matrix gene ontology groups, which are known to be central to skeletal muscle remodelling and gain in physiological capacity. Thus, despite the lack of change in global protein synthesis, the types of protein being synthesised are most probably extremely different in the skeletal muscle of ICU patients. As we wish to intervene to prevent the loss of muscle tissue function, it is important to determine which biological pathways were regulating this shift in muscle phenotype. To examine this question, we took a number of approaches. Using a multiple array analysis strategy we utilised the extensive animal models of muscle wasting, inactivity and inflammation from the Goldberg laboratory. While it is implausible to directly assess muscle function in the ICU setting, 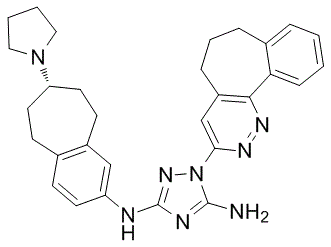 this comparative analysis allows us to contrast ICU patients with models of muscle atrophy, muscle inflammatory and muscle inactivity along the lines presented by Sacheck et al. It also facilitates discussion of the utility of such models and whether they accurately represent human muscle wasting disorders. In two recent publications, common genes up regulated or down regulated in animal models with muscle wasting have been identified. In the first study animal models for fasting, cancer cachexia, uremia and diabetes mellitus identified 120 unique genes involved in catabolism, and the authors named these ‘atrogens’. In a second paper these genes were Butenafine hydrochloride compared with two animal models of disuse induced muscle wasting, leading to 53 commonly changed genes.
this comparative analysis allows us to contrast ICU patients with models of muscle atrophy, muscle inflammatory and muscle inactivity along the lines presented by Sacheck et al. It also facilitates discussion of the utility of such models and whether they accurately represent human muscle wasting disorders. In two recent publications, common genes up regulated or down regulated in animal models with muscle wasting have been identified. In the first study animal models for fasting, cancer cachexia, uremia and diabetes mellitus identified 120 unique genes involved in catabolism, and the authors named these ‘atrogens’. In a second paper these genes were Butenafine hydrochloride compared with two animal models of disuse induced muscle wasting, leading to 53 commonly changed genes.
To capture a broad assessment of the molecular processes represented by the modulated genes
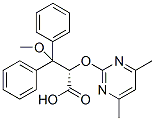
Leave a reply
 the aggregation of mutant huntingtin or ataxin-3 proceeds by nucleated growth polymerization into protein fibrils that structurally resemble amyloid fibrils and react with amyloid-specific histochemical dyes such as Congo Red or thioflavin. Analogous biophysical properties are observed for b-amyloid and a-synuclein fibrils associated with Alzheimer��s and Parkinson��s diseases, respectively. Amyloid fibrils are thought to be nucleated by monomers or globular oligomers of misfolded protein. In turn, fibrils may co-assemble into much larger insoluble protein aggregates that are resistant to proteolysis. While it is clear that protein misfolding can elicit cellular toxicity, whether fibrillar protein aggregates are
the aggregation of mutant huntingtin or ataxin-3 proceeds by nucleated growth polymerization into protein fibrils that structurally resemble amyloid fibrils and react with amyloid-specific histochemical dyes such as Congo Red or thioflavin. Analogous biophysical properties are observed for b-amyloid and a-synuclein fibrils associated with Alzheimer��s and Parkinson��s diseases, respectively. Amyloid fibrils are thought to be nucleated by monomers or globular oligomers of misfolded protein. In turn, fibrils may co-assemble into much larger insoluble protein aggregates that are resistant to proteolysis. While it is clear that protein misfolding can elicit cellular toxicity, whether fibrillar protein aggregates are 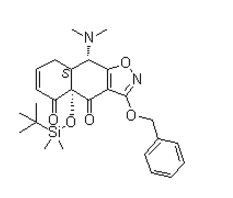 of the transporter since our results indicate that the triple mutation DDD of 611, 613, and 616 retained only 5% of its 5HT uptake capacity as compared to control SERT. It is also important to note that the presence of such a large amount of negative charge on the end of the protein could cause alterations in protein folding or proteinprotein associations that are important for protein function, resulting in the observed blunting of transport capacity. Next, we analyzed the impact of four truncations of the SERT C-terminus on the trafficking and expression of SERT on the plasma membrane using biotinylation and IF assays. Our data indicate that depending on the amount of truncation from the Cterminus of SERT, there was altered localization of the transporter. Therefore, we carried out a biotinylation analysis on some of the phosphorylation-mimicking mutations in an effort to determine the plasma membrane localization of these mutants, i.e., whether the mutation arrests them intracellularly or whether the mutants can still traffic to the plasma membrane. The data indicate that 3 possible phosphorylation sites do contribute to the 5HT uptake rates of transporters via inhibiting their proceedings toward the plasma membrane.
of the transporter since our results indicate that the triple mutation DDD of 611, 613, and 616 retained only 5% of its 5HT uptake capacity as compared to control SERT. It is also important to note that the presence of such a large amount of negative charge on the end of the protein could cause alterations in protein folding or proteinprotein associations that are important for protein function, resulting in the observed blunting of transport capacity. Next, we analyzed the impact of four truncations of the SERT C-terminus on the trafficking and expression of SERT on the plasma membrane using biotinylation and IF assays. Our data indicate that depending on the amount of truncation from the Cterminus of SERT, there was altered localization of the transporter. Therefore, we carried out a biotinylation analysis on some of the phosphorylation-mimicking mutations in an effort to determine the plasma membrane localization of these mutants, i.e., whether the mutation arrests them intracellularly or whether the mutants can still traffic to the plasma membrane. The data indicate that 3 possible phosphorylation sites do contribute to the 5HT uptake rates of transporters via inhibiting their proceedings toward the plasma membrane.