Though it has been applied to studies of cultivated plants and to a lesser extent, conifers. Our efforts permit parameter estimation for biologically meaningful demographic models and provide a direct measure of our confidence in the model and its relevance to our data. Our results build on previous work that documents high differentiation among A. lyrata populations, and points to central European populations as a center of diversity for A. lyrata ssp. petraea. Other studies have further argued that central European populations may have served as refugia from which Northern Europe was re-colonized after glacial cycles during the Pleistocene, and even specifically hypothesized that the Icelandic population of A. lyrata ssp. petraea and North American populations of A. lyrata ssp. lyrata were colonized from Europe. Our results broadly concur with these ideas. Relative to the Central European population surveyed here, other populations reveal the hallmarks of population bottlenecks: lower diversity, loss of singleton and low frequency variants, higher LD and lower estimated r values. The demographic inferences summarized in Table 1 suggest strong bottlenecks with little subsequent recovery of size in the non-German populations. Moreover, although most loci show strong genetic structure, differentiation is lower with the German population. Pairwise comparisons also reveal a high proportion of shared variants and few fixed Butenafine hydrochloride differences between Germany and other populations. Even populations as different genetically and geographically as Canada and Russia each possess extensive shared variation with Germany, suggesting that the nonGerman populations sampled represent subsets of the diversity in Germany. Consistent with this, all of our pairwise comparisons show a higher proportion of unique variants in Germany. Both FST and Bayesian cluster analyses reveal unusually strong population structure for an outcrossing herbaceous species, providing little evidence for recent admixture or gene flow, but suggesting long-term persistence of isolated populations. This finding is supported by analysis of an alternate demographic model that explicitly estimated low pairwise migration between Germany and other populations. It is possible, of course, that migration from unsampled populations or species contributes to observed patterns of diversity. One would expect such migration to increase both diversity and LD, but our data show higher LD only in non-German populations with lower levels of diversity. Although the data to explicitly test this hypothesis are not currently available, our sequence data provide no compelling evidence that migration from unsampled populations has strongly affected our sampled populations. Although our demographic model does not aim to infer a definitive history, it is important to consider how inclusion of nonequilibrium processes may affect estimation of divergence times. Our estimates are much lower than calculations based solely on median pairwise FST values, which yields divergence times ranging from,90,000 years between Germany and Iceland to,170,000 years between Germany and Russia. However, our estimates are considerably older than the end of the most recent Ice Age, when Northern Europe was most likely re-colonized by A. lyrata. We note, however, that the 95% credible intervals of our estimates generally include times as recent as 10,000 years ago, and that because tS estimates in 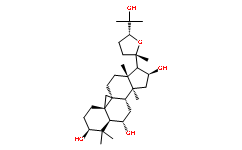 years are Chloroquine Phosphate proportional to the mutation rate, a rate twice as high as that estimated by Koch et al. would reduce the value in years of our divergence time.
years are Chloroquine Phosphate proportional to the mutation rate, a rate twice as high as that estimated by Koch et al. would reduce the value in years of our divergence time.
Explicit modeling of natural population history remains rare in studies of flowering plants
Leave a reply
 microarrays and computationally deconvolved into its ingredient cell lines. So although there appears to be systematic error, it is relatively small and not necessarily explained by the cell type. This characterization of performance on a test data set designed to simulate the challenges of deconvolving leukocytes provides important knowledge of the capabilities of the method that guide its application to whole blood. ummarized in Table 1. We selected probesets to use as the basis of discriminating between cell types by screening for those that
microarrays and computationally deconvolved into its ingredient cell lines. So although there appears to be systematic error, it is relatively small and not necessarily explained by the cell type. This characterization of performance on a test data set designed to simulate the challenges of deconvolving leukocytes provides important knowledge of the capabilities of the method that guide its application to whole blood. ummarized in Table 1. We selected probesets to use as the basis of discriminating between cell types by screening for those that  Cup, Hrb27C, and Smaug, as sumoylation targets, which have essential roles in the regulation of oskar mRNA localization and translation. This interesting and novel finding suggests a role of SUMO in regulating the functions of maternal mRNA by modifying components of oskar mRNP, and therefore could explain some of the pleiotropic defects observed in the embryonic patterning of embryos resulting from sumo mutant GLCs. The oskar mRNP is one of several instances in which
Cup, Hrb27C, and Smaug, as sumoylation targets, which have essential roles in the regulation of oskar mRNA localization and translation. This interesting and novel finding suggests a role of SUMO in regulating the functions of maternal mRNA by modifying components of oskar mRNP, and therefore could explain some of the pleiotropic defects observed in the embryonic patterning of embryos resulting from sumo mutant GLCs. The oskar mRNP is one of several instances in which 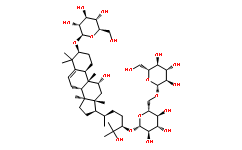 in a specific cell type, indicating a specific nature or physiologic function of each cell type. For instance, at least six proteins including TUBB2A protein, KRT8 protein, a-enolase, 14-3-3 protein sigma, HSP60, and myosin-9 were expressed at significantly higher levels only in prES cells. Tubulins are the major components of the filamentous structure of cellular microtubules with a-tubulin being the most common one. The microtubule plays many crucial roles in intracellular transport, cell morphology, polarity, signaling, and division of the cell, which also make it a target for the study of cancer therapy. In this study, we found that a-tubulin or tubulin-b was upregulated in both f-rES and p-rES cells, strongly suggesting that ES cells are one of the actively proliferating cell types compared to the terminally differentiated fibroblasts. Moreover, previous studies have also shown that a-tubulins in mES cells are downregulated along with vimentin, one of the intermediate
in a specific cell type, indicating a specific nature or physiologic function of each cell type. For instance, at least six proteins including TUBB2A protein, KRT8 protein, a-enolase, 14-3-3 protein sigma, HSP60, and myosin-9 were expressed at significantly higher levels only in prES cells. Tubulins are the major components of the filamentous structure of cellular microtubules with a-tubulin being the most common one. The microtubule plays many crucial roles in intracellular transport, cell morphology, polarity, signaling, and division of the cell, which also make it a target for the study of cancer therapy. In this study, we found that a-tubulin or tubulin-b was upregulated in both f-rES and p-rES cells, strongly suggesting that ES cells are one of the actively proliferating cell types compared to the terminally differentiated fibroblasts. Moreover, previous studies have also shown that a-tubulins in mES cells are downregulated along with vimentin, one of the intermediate